
In humans, ciliary muscles contract to change the tension of zonule fibers to deform the lens, leading to an increase in the lens’ optical power to focus on nearby objects, a process known as accommodationThis image shows a 3D reconstruction of a confocal z-stack through a human lens with attached cilliary body and zonules. The zonules are stained with wheat germ agglutinin (cell membrane marker, red) and microfibril-associated glycoprotein MAGP-1 (zonules, yellow). The ciliary body (in the foreground of the image) and the lens (behind the zonules) are stained for filamentous actin (F-actin, phalloidin, green) and cell nuclei (DAPI, blue).
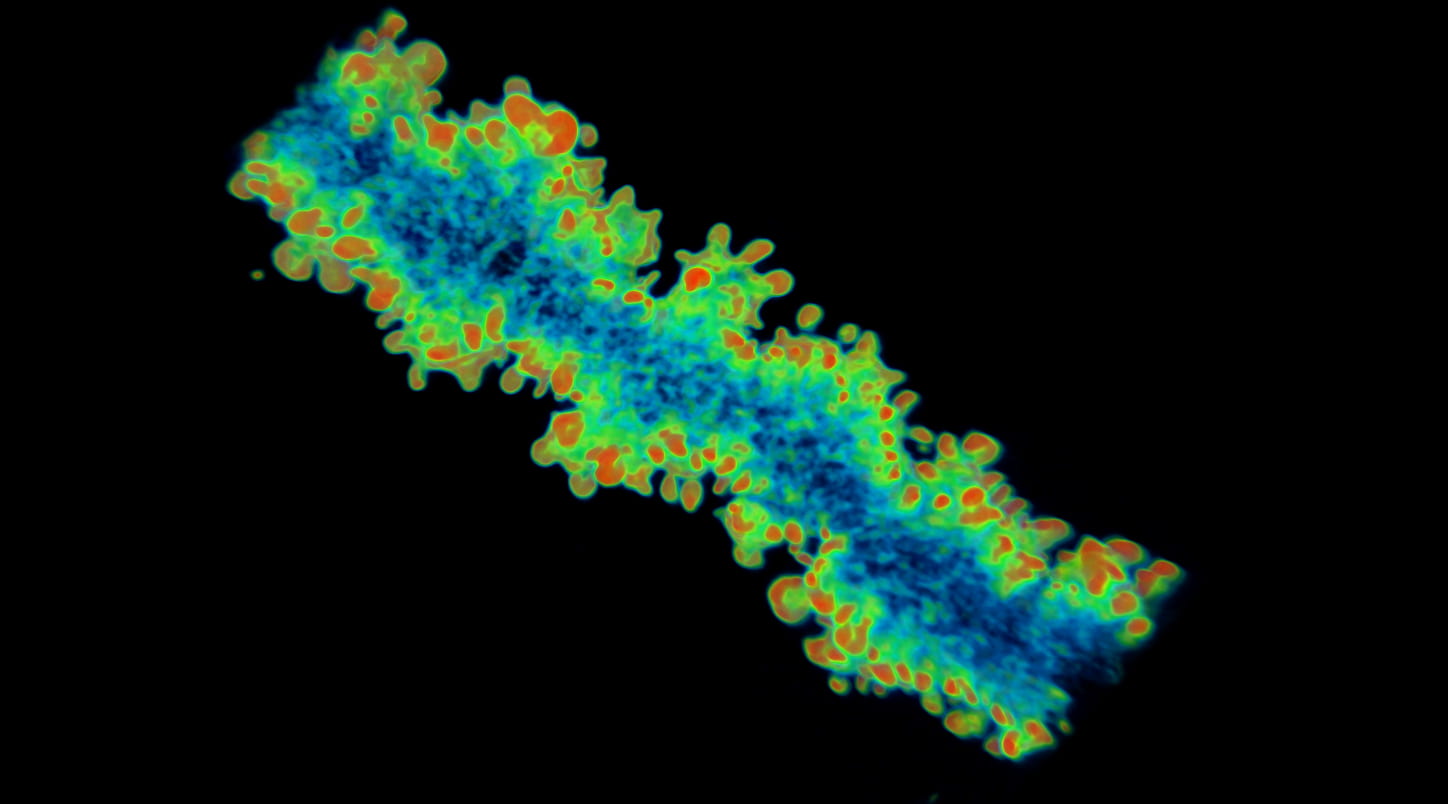
Eye lens fiber cells have complex membrane interdigitations that are required for biomechanical properties. This 3D reconstruction of a single lens fiber stained with phalloidin for F-actin (staining signal is color coded from red-purple for 255-0 arbitrary intensity units) showing that interdigitations have a rich F-actin network. Actin-binding proteins, including tropomyosin 3.5 and tropomodulin 1, are required for select F-actin networks required for supporting lens fiber cell biomechanical properties. For more information, please see Dr Cheng’s article in JCS.
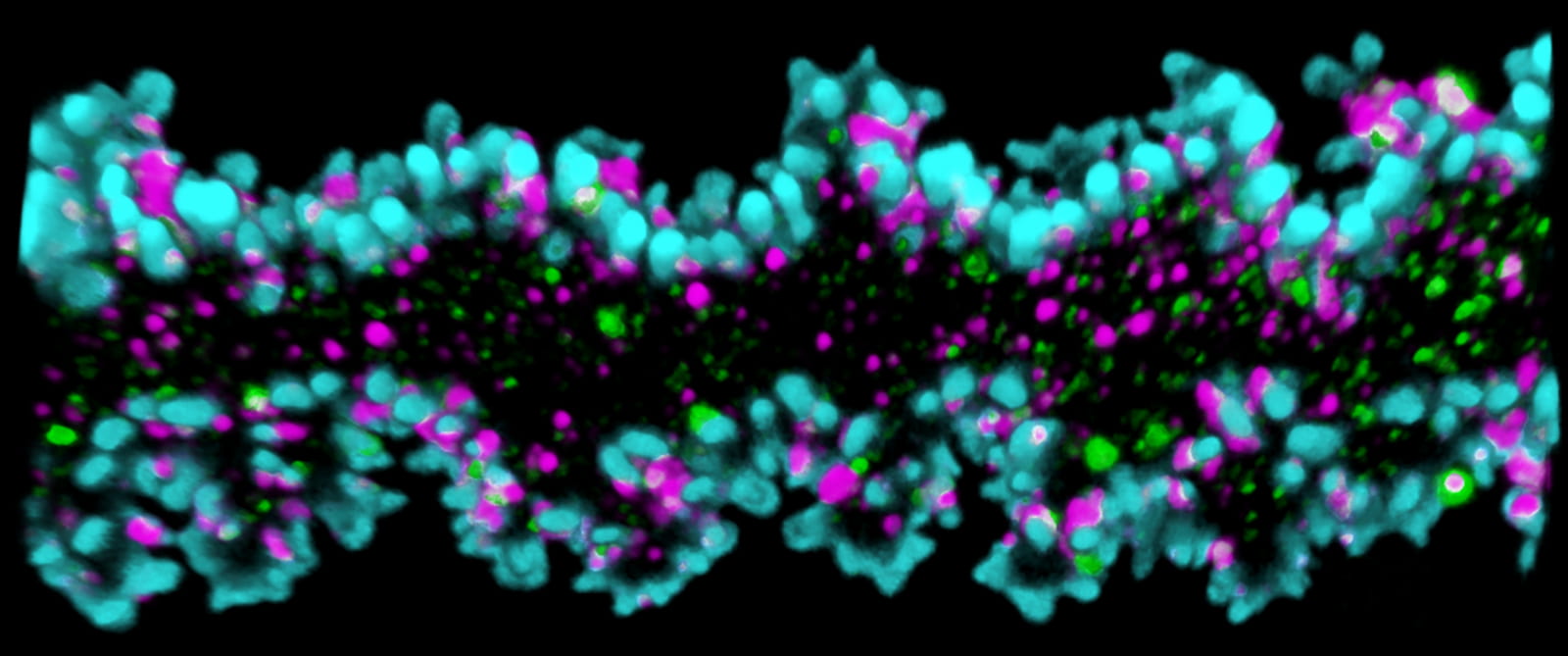
This 3D reconstruction of a single mouse lens fiber cell stained with phalloidin for F-actin (teal). Actin-binding proteins, including tropomyosin 3.5 (magenta) and tropomodulin 1 (green), support select F-actin networks required for normal lens fiber cell morphology and biomechanical properties. For more information, please see Dr Cheng’s article in IOVS.
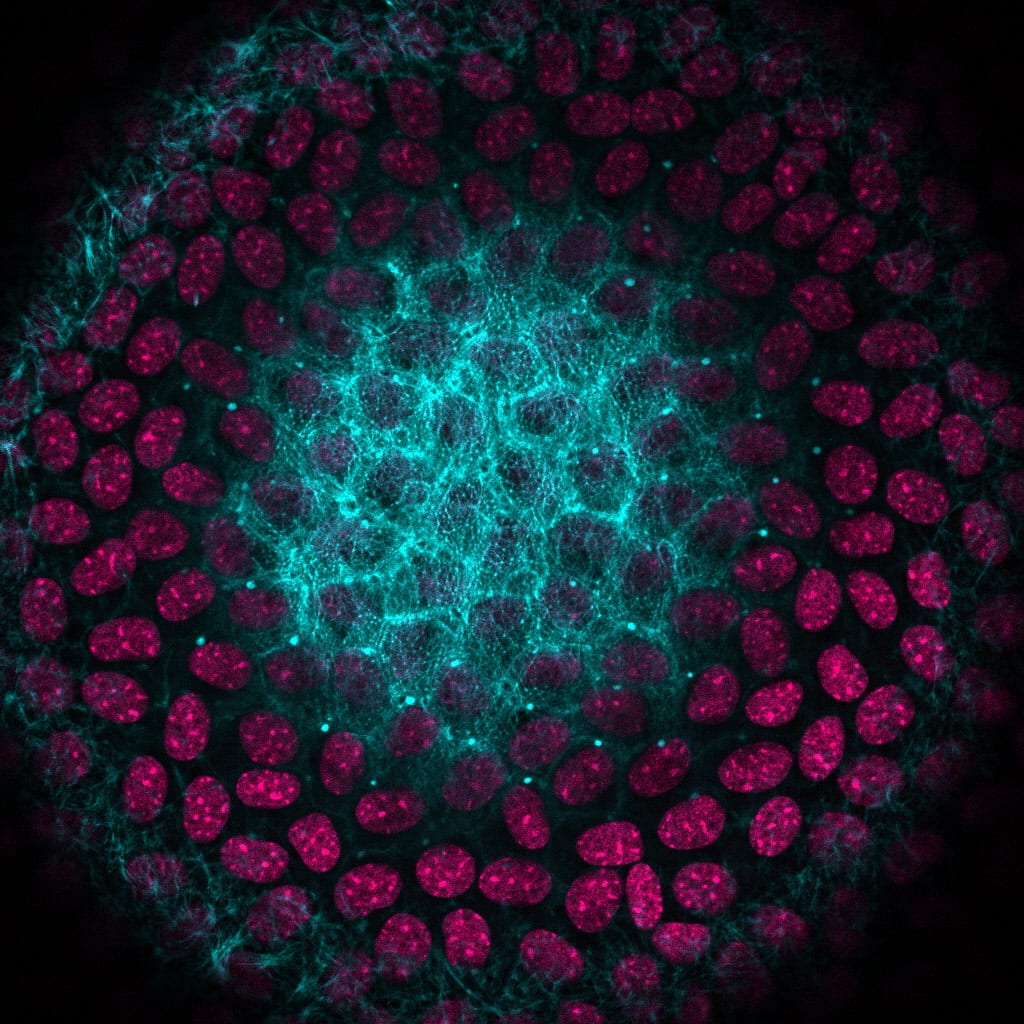
A monolayer of epithelial cells covers the anterior hemisphere of the lens. Epithelial cells near the anterior pole are cuboidal in shape and contain diverse actin filaments. This image shows one optical section of a whole mouse lens stained for F-actin (phalloidin, teal) and cell nuclei (DAPI, magenta). The basal surfaces of these anterior epithelial cells contain actin stress fibers and actin filament-rich lamellipodia (cells in the outermost ring). On the lateral surfaces, anterior epithelial cells are polygonal in shape with actin cortical fibers and sequestered actin bundles (cells in second ring from the outside with bright teal F-actin puncta). Apical domains of the anterior epithelial cells contain polygonal arrays of actin filaments (cells in the middle). These geodesic domes of actin are hypothesized to flatten in order to maintain epithelial cell integrity during lens shape changes (accommodation) so you can see near objects clearly.
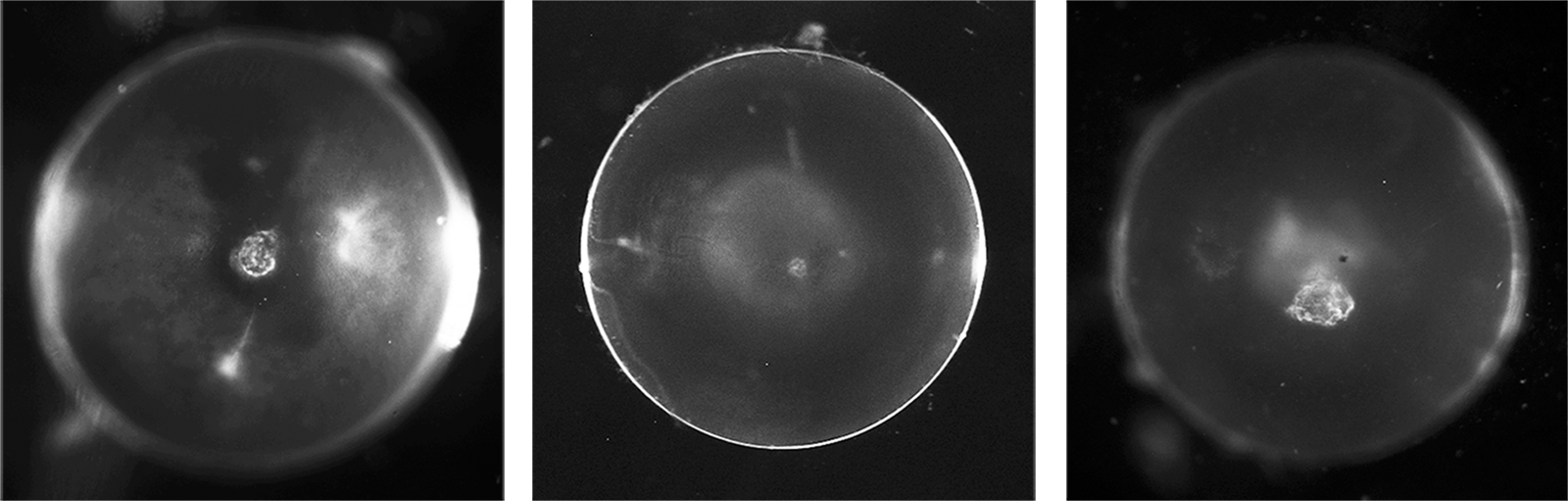
Photos of a wild-type (transparent lens on the left) and lenses with anterior (middle) or nuclear (right) cataracts. Scale bar, 1mm. For more information in Eph-ephrin bidirectional signaling in the lens, please check out the following articles:
EphA2 and ephrin-A5 are not a receptor-ligand pair in the ocular lens.
EphA2 and Src regulate equatorial cell morphogenesis during lens development.
Diverse roles of Eph/ephrin signaling in the mouse lens.

Life-long lens growth requires the differentiation, migration and elongation of equatorial epithelial cells into secondary fiber cells. Secondary fiber cells elongate toward the anterior and posterior poles and meet at the anterior and posterior lens sutures. This image shows the anterior suture of a mouse lens where the tips of elongating fiber cells meet at the pole (en face view). The fiber cells are stained for F-actin (phalloidin, grayscale), which shows that actin filaments are enriched at fiber cell tips. For more information on the actin cytoskeleton in the lens, please see Dr Cheng’s review in Experimental Eye Research.